
Меню сайта
Наши новости
Распространение алкалоидов в растительном мире.
Умягчение воды
Механизмы гомогенного катализа.Статьи и работы по химии / Гомогенный катализ / Статьи и работы по химии / Гомогенный катализ / Механизмы гомогенного катализа. Механизмы гомогенного катализа.Страница 3
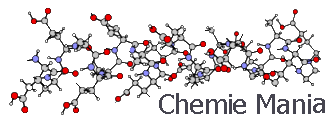
Рис.
1. Схема активного центра фермента карбоксипептидазы (по данным Липс-комба, Рика, Хартсака, Кешо и Бетджа):
Показаны фрагменты пептидном цепи с функциональными боковыми группами. Цифры обозначают порядковые номера остатков аминокислот, которым принадлежат эти функциональные группы. Молекула субстрата изображена с утолщенными связями. В шести-членном активированном комплексе штрихами показаны образующиеся связи, л сплошными линиями — разрывающиеся связи
В результате этих взаимодействий, которые закрепляют в двух точках С-концевой остаток субстрата, пептидная связь в случае, если С-концевая аминокислота представляет собой L-изомер, оказывается направленной на каталитический центр фермента, представленный ионом цинка и оксигруппой тирозина. Поляризация связи С=О ионом цинка облегчает нуклеофильную атаку молекулы воды на электрофильный атом С. Участие оксигруппы тирозина обеспечивает синхронное протекание разрыва трех связей и образования трех новых связей в циклическом шести центровом активированном комплексе.
На этом примере видны некоторые важнейшие черты, свойственные большому числу ферментов. Во-первых, катализатор имеет как бы два центра — связывающий (контактный) и собственно каталитический. Один из них, представленный в рассмотренном случае протонированной гуанидиновой группой и тремя гидрофобными радикалами, обеспечивает образование комплекса фермент — субстрат (связывание субстрата ферментом), в результате чего расщепляемая связь направляется на каталитический центр. Собственно каталитический центр представлен в рассмотренном случае ионом цинка и оксигруппой тирозина.
Во-вторых, на этом примере видны структурные основы высокой специфичности ферментов, в частности стереоспецифичности. Так, если бы С-концевая аминокислота была о-изомером, то в рассматриваемом случае в сторону каталитического центра оказался бы направленным атом Н, а не группа — NН—СО—, и каталитический процесс не смог бы произойти.
Из изложенного ясно также, почему фермент катализирует разрыв пептидной связи именно С-концевой аминокислоты и имеет преимущественное сродство к остаткам ароматических аминокислот. Действительно, именно взаимодействие заряда концевой карбоксильной группы и наличие гидрофобного ароматического остатка обеспечивает взаимодействие субстрата с контактным центром фермента, которое обеспечивает нужную ориентацию гидролизуемой связи относительно каталитического центра.
Смотрите также
Антибиотики
Антибиотики, вырабатываемые микроорганизмами химические вещества, которые способны тормозить рост и вызывать гибель бактерий и других микробов. Противомикробное действие антибиотиков имеет избирательн ...
Радиоактивность
...