
Меню сайта
Наши новости
Распространение алкалоидов в растительном мире.
Умягчение воды
Ферментативный катализУчим химию / Учим химию / Ферментативный катализ Ферментативный катализСтраница 5
Ro = k2[ES]
Приняв, что [Eo] – общая концентрация фермента, а ([Eo] - [ES]) соответствует концентрации свободного фермента, а также что [S] >> [Eo], можно получить выражение для [ES]:
[ES] = ([Eo]∙ [S]/{ [S] + (k2 + k-1)/k1}
Отношение (k2 + k-1)/k1 называется константой Михаэлиса ( КМ); с учетом этого концент-
рация фермент-субстратного комплекса и начальная скорость могут быть описаны уравне-ниями:
[ES] = [Eo]∙ [S]/ (КМ + [S])
Ro = k2[ES] = k2[Eo]∙ [S]/ (КМ + [S])
Последнее уравнение называют уравнением Михаэлиса-Ментен. Необходимо отметить, что величина КМ совпадает с термодинамической константой диссоциации фермент-субстратного комплекса только в случае квазиравновесия первой стадии и лимитирования процесса второй стадией. Во всех остальных случаях КМ является сложным комплексом констант скорости стадий ферментативного процесса.
Рассмотрим механизм функционирования ферментативного катализатора на примере гидролитического фермента химотрипсина.
Химотрипсин – фермент поджелудочной железы, функция которого в организме заключается в расщеплении белков пищи, т.е. пептидной связи. Кроме этого химотрипсин может катализировать гидролиз сложных эфиров и некоторые другие реакции. Брутто формула химотрипсина, включающего 241 остаток аминокислот, не несет информации о строении: С1105H1732O344N300S12, также как перечисление количества аминокислотных остатков: аланин22 аргинин3 аспарагиновая кислота8 аспарагин14 глутаминовая кислота3 глутамин10 глицин24 гистидин2 изолейцин10 лейцин19 лизин14 метионин2 полуцистин10 пролин9 серин28 треонин22 триптофан8 тирозин4 валин23 фенилаланин6. Перечисленные аминокислотные остатки соединены в полипептидную цепь в определенной последовательности (первичная структура). Отдельные части полипептидной цепи за счет образования дополнительных связей (см.выше) скручиваются в α-спирали, β-тяжи и петли (вторичная структура). Перечисленные элементы вторичной структуры за счет дополнительных взаимодействий сворачиваются в два домена, в месте соприкосновения которых возникает активный центр фермента, включающий остаток серина (Х – -СН2ОН ), аспарагиновой кислоты ( Х - -СН2СОО-), гистидина.
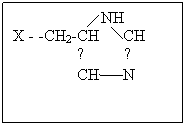
Механизм реакции гидролиза сложного эфира показан на схеме. 2. При подходе субстрата к активному центру фермента неполярная гидрофобная часть субстрата взаимодействует с гидрофобной частью активного центра, протон от серина переходит на азот гистидина, а протон от второго азота гистидина смещается к аниону остатка аспарагиновой кислоты. Образовавшийся из гидроксильной группы серина сильный нуклеофил - -О атакует электрофильный углерод субстрата, в то время как нуклеофильная часть субстрата взаимодействует с протоном, связанным с гистидином. В результате этих взаимодействий образуется фермент-субстратный комплекс. На следующей стадии рвется связь С-Х в субстрате, уходит молекула НХ, а ее место в активном центре занимает молекула воды. Протон от остатка аспарагиновой кислоты возвращается к второму азоту гистидина. Затем рвется предварительно активированная связь О-Н в молекуле воды (протон связывается с первым азотом гистидина, а гидроксил – с углеродом бывшего субстрата). Протон от второго азота гистидина опять возвращается к остатку аспарагиновой кислоты. И наконец выделяется кислота, место которой занимает новый субстрат или активный центр возвращается в исходное состояние.
Смотрите также
Синтез и свойства комплексов железа (II) и железа (III)
Цель данной работы состоит в изучении строения и свойств комплексных
соединений железа (II) и железа (III).
В ходе выполнения работы были поставлены следующие задачи:
1) изучение литер ...
Применение каучуков
Фторкаучуки – каучуки
специального назначения, применяемые в производстве изделий и деталей, в
которых должны сочетаться высокая стойкость к действию агрессивных сред и
теплостойкость. Фторкаучуки ...
Синтез 2,2-диэтоксииндандиона
Аминокислоты, пептиды
и протеины, или белки образуют группу химически и биологически родственных
соединений, которым принадлежит очень важная роль в жизненных процессах. При
полном гидролиз ...